有糸分裂とは何ですか?
有糸分裂は、同じ遺伝情報を持つ2つの娘核をもたらす単一細胞核の分裂です。 それは真核生物の細胞で起こります-原核生物は細胞核を持っていません-そして通常、2つの同一の娘細胞が出現する細胞全体の分裂に先行します。
真核細胞の分裂の細胞周期では、核分裂と細胞分裂が結びついています。 したがって、有糸分裂と細胞質分裂は、まとめて有糸分裂またはM期とも呼ばれます。 連続する有糸分裂の間期の間、 DNA染色体の分子が2倍になり(複製)、その後、各染色体は2つの同一の姉妹染色分体で構成されます。 有糸分裂の間、これらの染色分体は分離されて分割され、各娘核は娘染色体と同じ半分を受け取ります。 これは、母細胞の染色体ゲノム全体の同一のコピーを2つの娘細胞に渡すことができることを意味します。
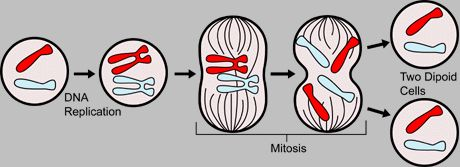
有糸分裂では、染色体の数に変化はなく、倍数性の程度は同じままです。 親細胞が一倍体であった場合、娘細胞の核も一倍体です。 最初の細胞が二倍体であった場合、娘細胞の核も二倍体です。
有糸分裂は主に多細胞生物の体の体細胞で起こりますが、生殖細胞の細胞分裂のタイプは減数分裂と呼ばれます。 単細胞生物の場合、有糸分裂は無性生殖の一形態と見なすことができます。
減数分裂は、姉妹染色分体が最初の細胞分裂で分離されないが、娘核に相同染色体として一緒に割り当てられる、根本的に異なる核分裂の方法によって有糸分裂と区別することができます。 それは世代サイクルに統合され、染色体セットと遺伝的に多様な娘細胞の減少につながります。
Christina Swords、Ph.D。が編集
有糸分裂の機能
有糸分裂は、2つの娘細胞核が同じ遺伝情報を再び受け取るように染色体に含まれる遺伝情報を分割することを可能にします。 これが起こるためには、母細胞の核内の遺伝物質が最初に複製されていなければなりません-細胞周期の前の間期の間に。 核分裂後の最初の1つの染色分体で構成される各染色体には、セントロメアで接続された2倍化後の2つの同一の姉妹染色分体があります。 有糸分裂段階では、これらは圧縮され、付着され、配置され、分離され、離れて移動されるため、空間的に異なる2つの(ただし染色体の数と種類は同じ)順序付けられたコレクションが形成され、その間で核が分割されます。

多細胞真核生物では、有糸分裂は新しい細胞核の形成の前提条件であり、通常は(いくつかの例外を除いて)新しい細胞の形成の前提条件です。 ヒトなどの多細胞生物では、発生中にすべての発生細胞株で細胞分裂が起こるわけではありません。 したがって、分化が完了すると、神経細胞と筋肉細胞は増殖しません。 これらの細胞は有糸分裂後に分裂周期を離れ、いわゆるG0期に入るので、DNAはまったく複製されません。 成熟したヒト赤血球は、細胞核を欠き、有糸分裂を開始できないため、分裂できなくなります。 一方、腸と皮膚の上皮細胞は、平均よりもはるかに頻繁に増殖するため、体の内面と外面を再生します。
ヒト細胞の実際の核分裂には通常約1時間かかります。有糸分裂期の間に起こる、連続的に分裂する細胞の細胞周期の間期は、細胞の種類に応じて、かなり長く、約12〜24時間続きます。 他の生物では、有糸分裂の持続時間は、ソラマメのように約2時間長くなることもあれば、ミバエのように短くなることもあります。
有糸分裂は、マイトジェンと呼ばれるさまざまなペプチドまたはタンパク質によって刺激される可能性があります。 例としては、成熟促進因子(MPF)、キナーゼを含むサイクリンBのタンパク質構造(CDK 1)があります。
有糸分裂と減数分裂の違い
有糸分裂と区別するために、減数分裂は特殊なタイプの核分裂であり、染色体セットの減少が起こり、同一の娘核は形成されません。 これは、有性生殖のための性細胞(すなわち、卵細胞と精子細胞)の形成で発生し、2つの分裂段階で2倍体の開始細胞から4つの一倍体細胞の形成をもたらす可能性があります。
減数分裂iと減数分裂iiの2つの段階があります。 第1段階(還元分裂)では染色体セットが半分になり、第2段階(方程式分裂)は終期iiの追加段階を伴う有糸分裂の過程にほぼ対応します。
有糸分裂の段階
概要概要
有糸分裂が始まる前に、ゲノム全体がDNAポリメラーゼ。
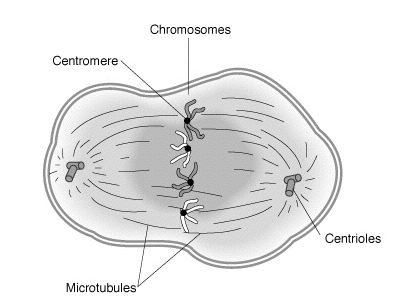
動物細胞の前期では、2つの中心体が分離し、細胞の反対の極に移動します。 中心体は微小管形成中心(MTOC)として機能し、それぞれが有糸分裂紡錘体アセンブリの開始点です。 高等植物では、他の細胞成分が中心体を持たないため、MTOCの役割を引き継ぎます。 染色体は凝縮し、光学顕微鏡で見えるようになり、今ではしばしば描かれるX字型でしか見えません(間期の間、染色体は細い糸のような構造として、長さ数センチまでの細長い形で存在します)。 染色体は間期ですでに複製されているので、それらはそれぞれセントロメアで接続されている2つの同一の姉妹染色分体で構成されています。 核膜が断片化すると、前期の終わりに到達します。
前中期では、核鞘が崩壊し、紡錘体装置の紡錘体繊維が両極から細胞の中心に向かって貫通します。 これで、付着した微小管を使って染色体を動かしたり、整列させたり、配置したりすることができます。
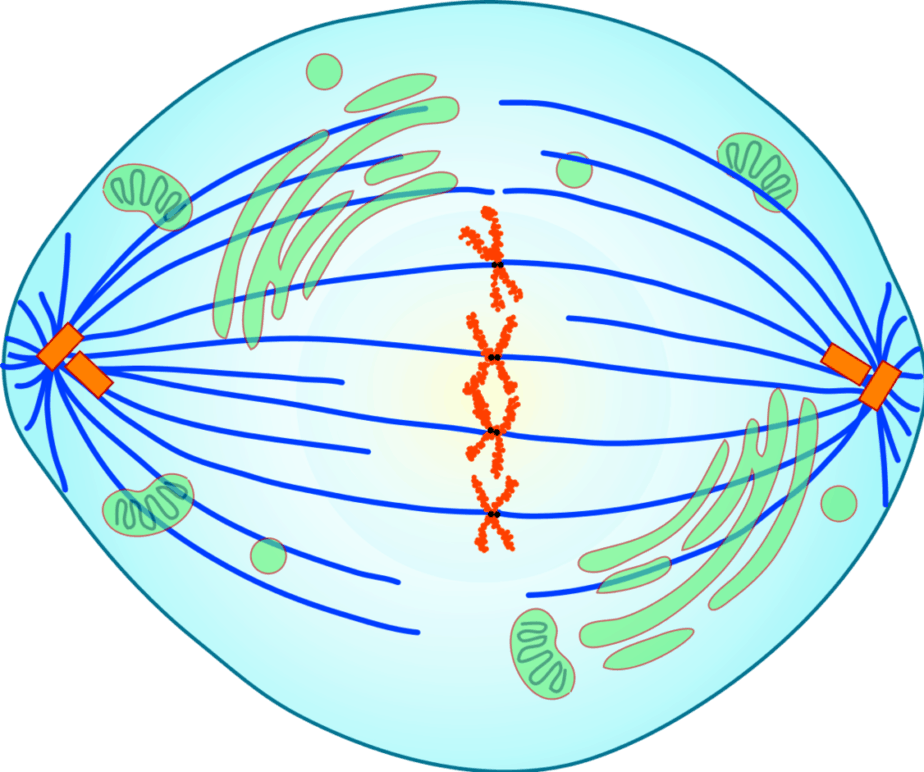
中期では、高度に凝縮した中期染色体が、細胞の中央にある紡錘体極間の紡錘体繊維として微小管によって整列されます。 染色体のすべてのペアがこの中期プレートに到着し、染色体が整列し、それらの動原体が両方の極からの微小管に接続されると、中期は完了します。
後期では、染色体の2つの染色分体が分離され、紡錘体繊維に沿って、最初はセントロメアで、紡錘体極に向かって反対方向に引き離されます。 このようにして、染色分体または娘染色体の完全なセットが各極で収集されます。 これにより、2つの娘核の基礎が作成されます。 後期は、2つの将来の娘核の染色体がそれ以上離れなくなったときに終了したと見なされます。
有糸分裂の最後の段階は終期と呼ばれます。 それは移行なしで前の後期に続きます。 動原体繊維が破壊され、核膜が再構築され、染色体が凝縮しなくなります。 脱凝縮が完了した後、遺伝子再び読み取ることができ、ニュークリアスは再び機能する形になります。
ほとんどの場合、終期の後に細胞質分裂が続き、それによって娘核を2つの娘細胞に割り当てることができます。 ただし、この細胞分裂は有糸分裂の一部ではありません。

前期
染色体凝縮
間期の間、染色体の連続したDNA二本鎖は、多くの部位でパッケージングタンパク質によって緩く囲まれているため、アクセス可能です。 前期の初めに、染色体を構成するクロマチンフィラメントは、ループ、コイル、およびダブルコイルの折り畳みと複数のターンによってコンデンシンを結合することにより、ますます凝縮および短縮します。
それらの高度にらせん状の構造のために、目に見える構造、染色体の核ループまたは染色分体が形成されます。 これらの新しい構造は、輸送に適したクロマチンフィラメントのよりコンパクトな形態を表しています。 また、この状態では、遺伝子のDNAセクションにアクセスできないため、発現できません。 このため、核小体は消えます。
紡錘繊維形成
動物細胞では、2つの中心体(それぞれが中心小体のペアを持つ)も間期の間に倍増することによって形成されています。 それらは今や核の反対側に移動し、したがって紡錘体の極を形成します。 中心体は、微小管から紡錘体装置の構造を組織化します。 最初に、紡錘繊維は星形の中心体から形成されます。これはアスターまたはアストラル微小管とも呼ばれます。
植物細胞は中心小体や中心体を使用しません。代わりに、他の構造が紡錘体装置の要素として微小管を組織化するタスクを引き継ぎます。
前中期
動物細胞では、前中期は核膜の分解から始まります。 中心体は反対の極に向かってさらに離れて押し出され、紡錘体繊維が伸びます。 発芽する有糸分裂紡錘体は、両極から核質に浸透し、極性微小管と呼ばれる極間の接続が重なり合っています。 紡錘繊維は染色体のセントロメアに付着し、動原体を形成します。 これらは、染色体の移動と整列、およびそれに続くセントロメア領域での染色分体の分離を可能にします。
中期
前中期が別の段階と見なされる場合、中期は有糸分裂の第3段階です。
紡錘体装置は、紡錘体極までほぼ等しい距離で、細胞の中央に染色体を配置します。 したがって、染色体は、姉妹染色分体を引き離すことができる開始位置に並んでいます。
この配置は中期プレートとも呼ばれます。 この段階の顕微鏡画像は、核型を決定するために、染色体のセットの個々の染色体を視覚的に識別するために使用されます。
有糸分裂のチェックポイントもこの段階に分類されます。微小管が紡錘体の両極に付着した後でのみ、染色分体間の結合が解放されます。
後期
染色体の2つの染色分体は分離され、異なる方向に移動します。 したがって、姉妹染色分体は娘染色体(1染色分体染色体)になり、紡錘体繊維に沿って細胞の反対の極に輸送されます。 このプロセスでは、動原体繊維が短縮されます。 その間、極繊維の微小管が長くなり、極が互いに離れる原因となる可能性があります。
染色体の移動(後期I)と紡錘体極線維の移動(後期II)を区別することができます。
終期
娘染色体が最終的に紡錘体極に到達すると、ますます短くなる動原体繊維が大幅に分解します。 極繊維は、極が最大距離に達するまで最初はさらに長くなる可能性があり、その時点でスピンドル装置が溶解します。 娘核の核膜は現在、古い核膜の断片から大部分が構築されています。 染色体は再び凝縮します。 核小体もそれぞれの核に再出現します。
細胞質分裂
ほとんどの場合、細胞は核の形成が完了した後に分裂します。 動物細胞では、アクチン繊維の収縮リングが細胞質に溝を作り、細胞を2つの娘細胞に分割します。
細胞分裂を伴わない有糸分裂
有糸分裂の後に細胞質分裂が続かない場合があります。 多細胞動物では、組織の分化は、機能を担う細胞がもはや分裂しない高度に秩序化された関係につながる可能性があります。 たとえば、神経系組織では、ほとんどのネットワーク化されたニューロンは有糸分裂後であり、分裂することができません。 成熟した心筋細胞にも分裂する能力がありません。
あなたは有糸分裂についてもっと学ぶことができますここに。