What is mitosis?
Mitosis is the division of a single cell nucleus that results in two daughter nuclei with the same genetic information. It takes place in cells of eukaryotic organisms – prokaryotes do not have a cell nucleus – and usually precedes a division of the entire cell, from which two identical daughter cells emerge.
In the cell cycle of dividing eukaryotic cells, nuclear division and cell division are coupled together. Mitosis and cytokinesis are therefore also referred to together as mitosis or M phase. During the interphase between successive mitoses, the DNA molecule of a chromosome is doubled (replication), after which each chromosome consists of two identical sister chromatids. During mitosis, these chromatids are then separated and split so that each daughter nucleus receives an identical half as a daughter chromosome. This means that an identical copy of the entire chromosomal genome of the mother cell can be passed on to two daughter cells.
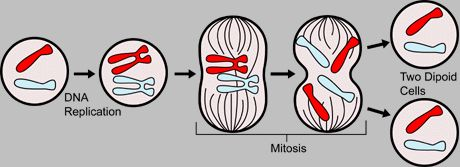
In mitosis, there is no change in the number of chromosomes, the degree of ploidy remains the same. If the parent cell was haploid, the nuclei of the daughter cells are also haploid. If the initial cell was diploid, the nuclei of the daughter cells are also diploid.
Mitosis mainly takes place in somatic cells in the body of multicellular organisms, while the types of cell division in reproductive cells are called meiosis. For single-celled organisms, mitosis can be considered a form of asexual reproduction.
Meiosis can be distinguished from mitosis by a fundamentally different way of nuclear division, in which the sister chromatids are not separated in the first cell division, but are assigned together as homologous chromosomes to a daughter nucleus. It is integrated into the generation cycle and leads to a reduction of the chromosome set and genetically diverse daughter cells.
Edited by Christina Swords, Ph.D.
Function of mitosis
Mitosis makes it possible to divide the genetic information contained in the chromosomes in such a way that two daughter cell nuclei receive the same genetic information again. For this to happen, the genetic material in the nucleus of a mother cell must first have been duplicated – during the preceding interphase of the cell cycle. Each chromosome, which initially consists of one chromatid after nuclear division, has two identical sister chromatids after doubling, which are connected at the centromere. In the mitosis phases, these are compressed, attached, arranged, separated and moved apart so that two spatially different – but identical in number and type of chromosomes – ordered collections are formed, between which the nucleus is then divided.

In multicellular eukaryotes, mitosis is the prerequisite for the formation of a new cell nucleus and usually – with a few exceptions – also for the formation of new cells. In multicellular organisms such as humans, cell division does not occur in all developed cell lines during their development. Thus, nerve cells and muscle cells do not multiply once differentiation is complete. These cells leave the division cycle post-mitotic and enter the so-called G0 phase, so that the DNA is not replicated at all. Mature human red blood cells can no longer divide because they then lack their cell nucleus and thus mitosis cannot be initiated. Epithelial cells in the intestine and skin, on the other hand, multiply much more frequently than the average and thus renew the inner and outer surfaces of the body.
The actual nuclear division of human cells usually takes about one hour; the interphase of the cell cycle of continuously dividing cells, which takes place between the mitosis phases, lasts considerably longer, about 12-24 hours, depending on the cell type. In other organisms, the duration of mitosis can be longer, as in the field bean with about two hours, or shorter, as in the fruit fly, where it is often only 9 minutes long.
Mitosis can be stimulated by various peptides or proteins called mitogens. An example is the maturation promoting factor (MPF), the protein structure of cyclin B with a kinase (CDK 1).
Difference between mitosis and meiosis
To distinguish from mitosis division, meiosis is a special type of nuclear division, in which a reduction of the chromosome set takes place and no identical daughter nuclei are formed. It occurs in the formation of sex cells (ie. egg cells and sperm cells) for sexual reproduction and can result in the formation of four haploid cells from a diploid starting cell in two division steps.
There are two stages: meiosis i and meiosis ii. In the first stage (reduction division) the chromosome set is halved, while the second stage (equation division) corresponds approximately to the course of mitosis with an extra stage of telophase ii.
Phases of a mitosis
Overview
Before mitosis begins, the whole genome is replicated by a DNA polymerase.
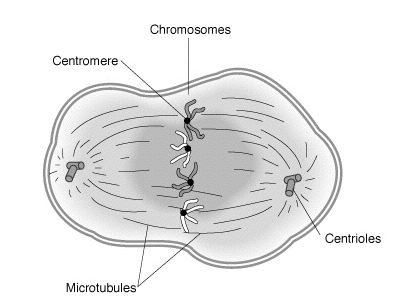
In the prophase of the animal cell, the two centrosomes separate and migrate to opposite poles of the cell. The centrosomes act as microtubule-organizing centers (MTOC) and are each starting points for mitosis spindle assembly. In higher plants, other cell components take over the task of MTOC, because their cells do not have centrosomes. The chromosomes condense, thus becoming visible under the light microscope, and are only now visible in the often depicted X-shape (during interphase they are present in an elongated form up to several centimeters long, as thin thread-like structures). Since the chromosomes have already been duplicated in the interphase, they consist of two identical sister chromatids each, which are still connected at the centromere. The end of prophase is reached when the nuclear envelope fragments.
In the prometaphase, the nuclear sheath disintegrates and the spindle fibres of the spindle apparatus penetrate from both poles towards the center of the cell. The chromosomes can now be moved, aligned and arranged by means of the adherent microtubules.
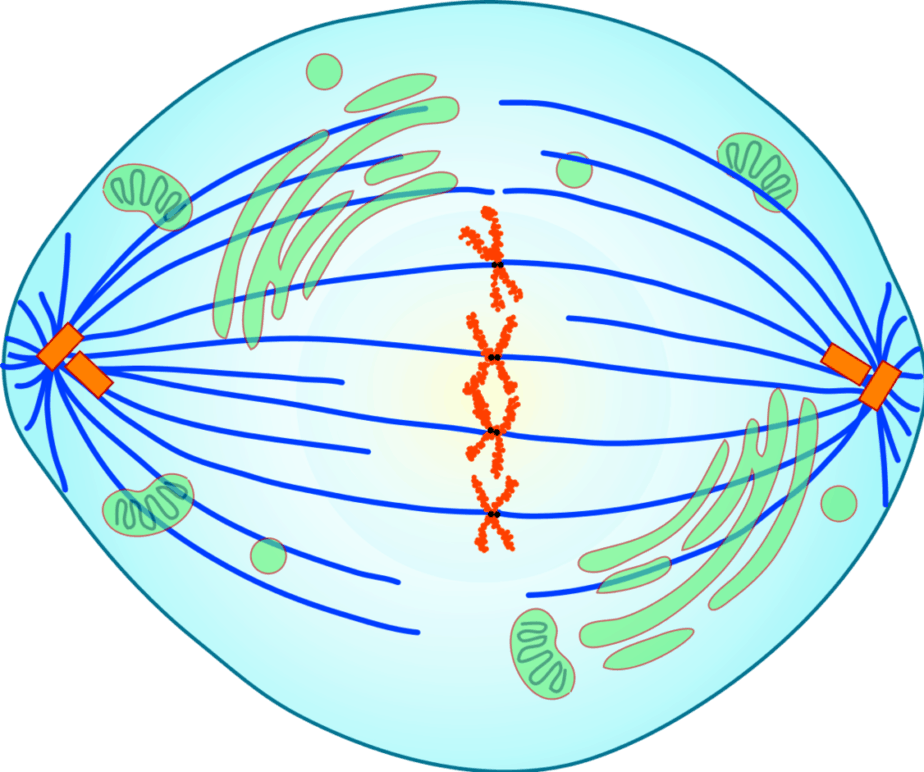
In the metaphase, the highly condensed metaphase chromosomes are aligned by the microtubules as spindle fibres between the spindle poles in the middle of the cell. The metaphase is complete when all pairs of chromosomes have arrived in this metaphase plate, chromosomes line up, and their kinetochores are connected with microtubules from both poles.
In the anaphase, the two chromatids of a chromosome are separated and pulled apart along the spindle fibers, centromere first, in opposite directions towards the spindle poles. In this way, a complete set of chromatids or daughter chromosomes is collected at each pole. This creates the basis for the two daughter nuclei. The anaphase is considered to be finished when the chromosomes of the two future daughter nuclei no longer move further apart.
The last phase of mitosis is called telophase. It follows the preceding anaphase without transition. The kinetochore fibres break down, the nuclear envelope is rebuilt and the chromosomes decondense. After the decondensation is completed, the genes can be read off again and the nucleus has its working form again.
In most cases, the telophase is followed by cytokinesis, with which the daughter nuclei can then be assigned to two daughter cells. However, this cell division is not part of mitosis.

Prophase
Chromosome condensation
During interphase, the continuous DNA double-strand of a chromosome is loosely surrounded by packaging proteins at many sites and is therefore accessible. At the beginning of the prophase, the chromatin filaments that make up a chromosome increasingly condense and shorten by binding condensins through folding and multiple turns in loops, coils, and double coils.
Due to their highly spiralized structure, visible structures are formed, the nuclear loops or chromatids of a chromosome. These new structures represent a more compact form of chromatin filaments suitable for transport. Also, in this state the DNA section of a gene is not accessible and thus cannot be expressed. Because of this, the nucleolus disappears.
Spindle Fiber Formation
In animal cells, two centrosomes (each with a pair of centrioles) have also been formed by doubling during interphase. They now migrate to opposite sides of the nucleus and thus form the poles of the spindle. The centrosomes organize the structure of the spindle apparatus from microtubules. At first, spindle fibres are formed from the centrosomes in a star shape, which is also called an aster or astral microtubule.
Plant cells do not use centrioles or centrosomes; instead, other structures take over the task of organising microtubules as elements of the spindle apparatus.
Prometaphase
In animal cells, the prometaphase begins with the degradation of the nuclear envelope. The centrosomes are pushed further apart towards opposite poles and the spindle fibers extend. The sprouting mitotic spindle penetrates from both poles into the nucleoplasm, with overlapping connections between the poles, called polar microtubules. Spindle fibers attach themselves at the centromeres of the chromosomes, forming kinetochores. These enable the movement and alignment of a chromosome and the subsequent separation of its chromatids in the centromere region.
Metaphase
The metaphase is the third phase of mitosis if the prometaphase is considered as a separate phase.
The spindle apparatus arranges the chromosomes in the middle of the cell, with approximately equal distance to the spindle poles. Thus, the chromosomes lie side by side in a starting position from which the sister chromatids can then be pulled apart.
This arrangement is also called the metaphase plate. Microscopic images of this phase are used to visually identify individual chromosomes of a set of chromosomes in order to determine the karyotype.
A checkpoint of mitosis also falls into this phase: only after microtubules have attached on both poles of the spindle can the binding between the chromatids be released.
Anaphase
The two chromatids of a chromosome are separated and moved in different directions. The sister chromatids thus become daughter chromosomes (one-chromatid chromosomes), which are transported along the spindle fibres to the opposite poles of the cell. In this process the kinetochore fibres are shortened. Meanwhile, the microtubules of the pole fibres can lengthen, causing the poles to move away from each other.
A distinction can be made between the moving apart of the chromosomes – as anaphase I – and the moving apart of the spindle pole fibers – as anaphase II.
Telophase
When the daughter chromosomes finally reach the spindle poles, the increasingly shortened kinetochore fibres break down to a large extent. The polar fibres can initially lengthen even further until the poles reach their maximum distance apart, at which point the spindle apparatus dissolves. The nuclear envelope of the daughter nuclei is now largely built up from fragments of the old nuclear membrane. The chromosomes decondense again. The nucleoli also reappear in each respective nucleus.
Cytokinesis
In most cases, the cell divides after the completion of the formation of the nucleus. In animal cells, a contractile ring of actin fibers furrows the cytoplasm, dividing the cell into two daughter cells.
Mitosis without cell division
Mitosis is sometimes not followed by cytokinesis. In multicellular animals, the differentiation of tissues can lead to highly ordered relationships in which function-bearing cells no longer divide. In nervous system tissue, for example, most networked neurons are post-mitotic and unable to divide. Mature heart muscle cells also have no ability to divide.
You can learn more about mitosis here.